Looking at Daniel Novotny’s Book (2013) “Ens Rationis from Suarez to Caramuel” (Part 7 of 19)
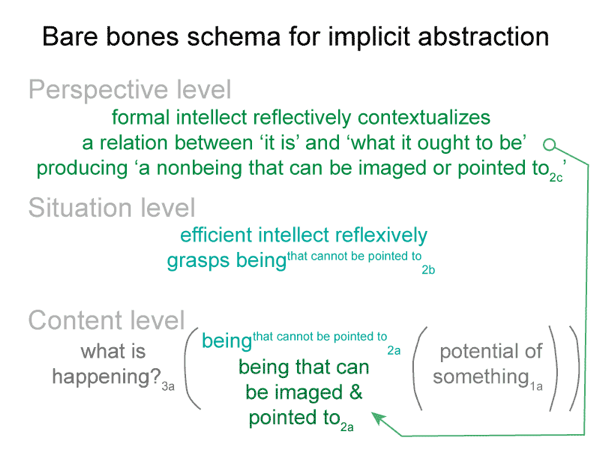
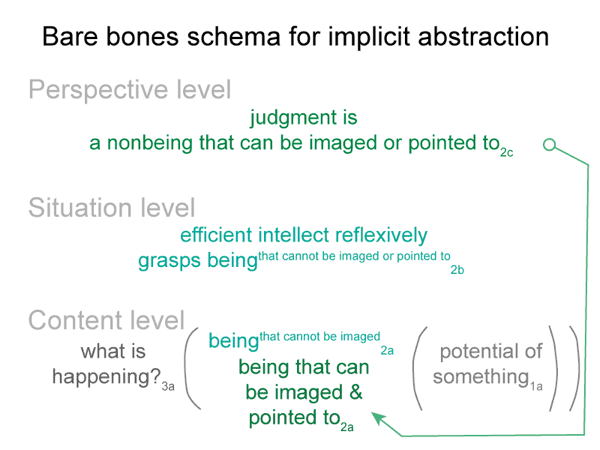
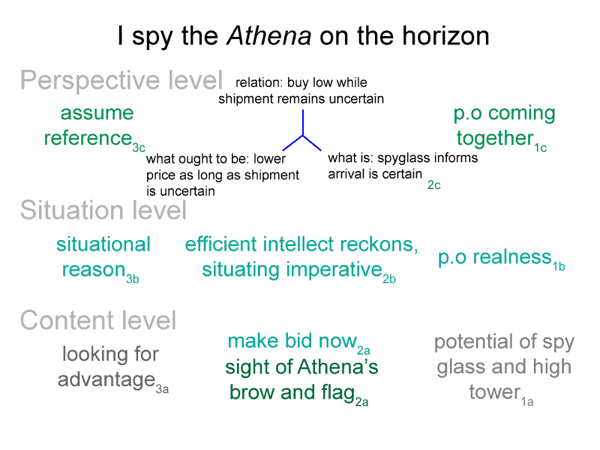
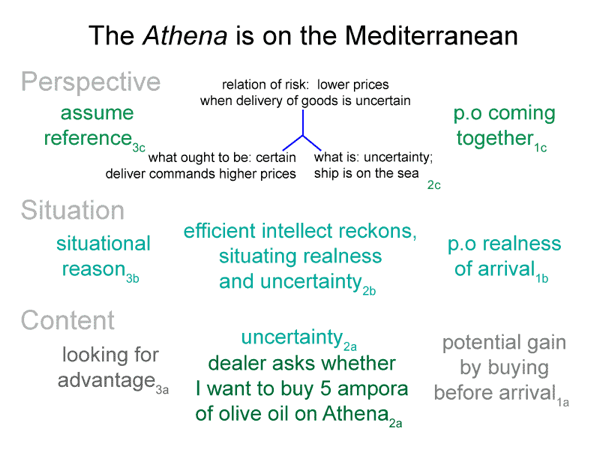
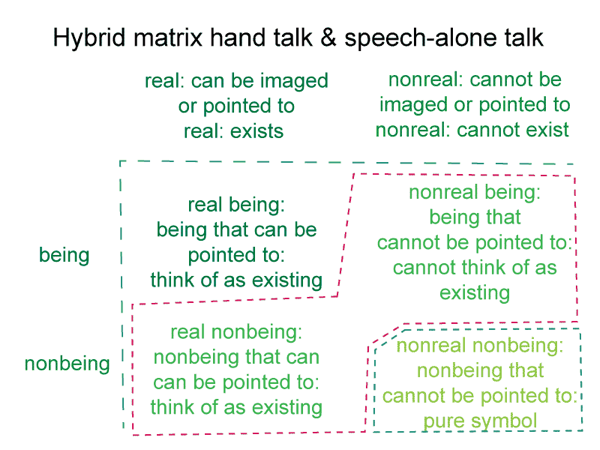
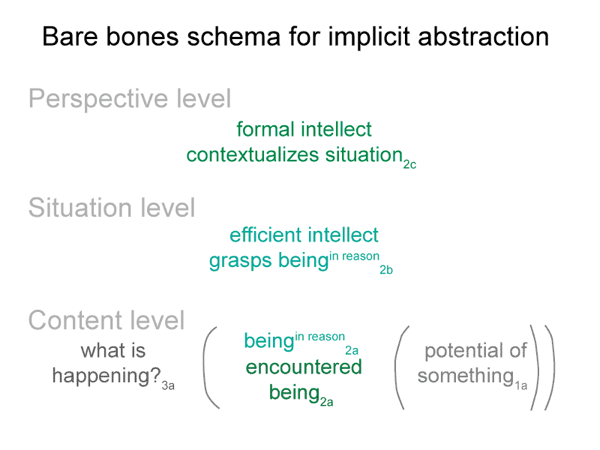
0062 Before I continue, however, let me show the bare-bones schema once again.
0063 Novotny lists Suarez’s nine claims about the nature of beings of reason.
SN1: ‘A being of reason’ is what cannot be actualized in reality.
What does this mean?
A beingin_reason cannot exist by itself.
0064 SN2: Not every being of reason is self-contradictory.
Why does Suarez mention this?
Consider the question that Suarez thought he was addressing: How do you explain negations, privations, self-contradictions and relations?
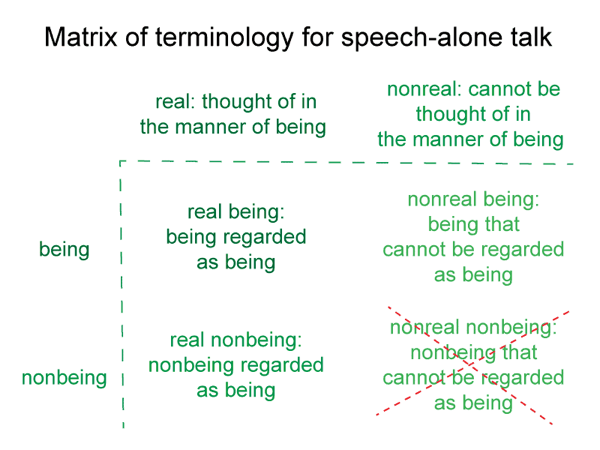
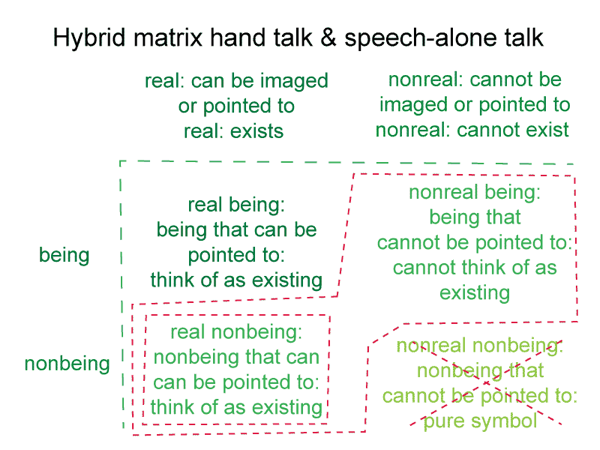
After all, they are nonexistent. They cannot exist by themselves. Yet, they can be real. One can think of them in the manner of being.
0065 Indeed, the model shows that the question that Suarez was addressing was more extensive and twisted than he imagined. Negations, privations, self-contradictions and relations are beings of reason2a. However, according to the model, they occur in conjunction with encountered beings2a.
0066 Here is an example. I dig with a shovel. The soil that I move goes with beingencountered. The hole2a goes with nonbeing. When I say, “Watch your step. Don’t fall into the hole that I just dug.” I speak of the hole as if it were a being. But, it is not a beingitself. It is a privation of soil.
If you fall into the hole, the hole itself does you no harm. It is the soil that you collide with that injures. The soil that you would hit in a fall corresponds to an encountered being2a.
0067 Anyone who has come across disturbed soil and has reflected on the types of injury produced by such encountershas a perspective on the danger of holes. That person reflexively grasps the danger of this nonbeing. It is as dangerous as any being that one might run into.
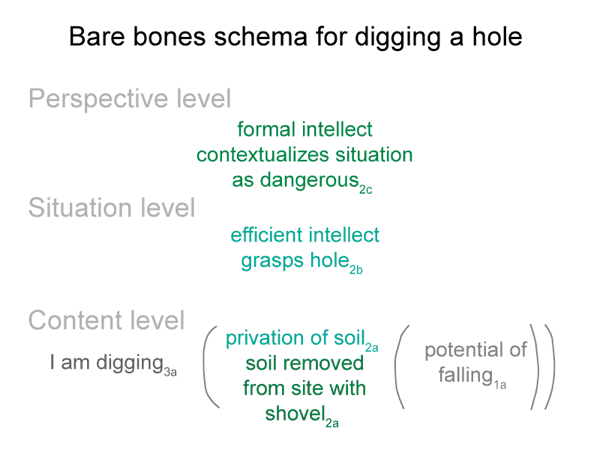
0068 A hole is a privation. A configuration of soil generates this being of reason. Privations are nonbeings that can be imagined as beings, but cannot exist in reality.
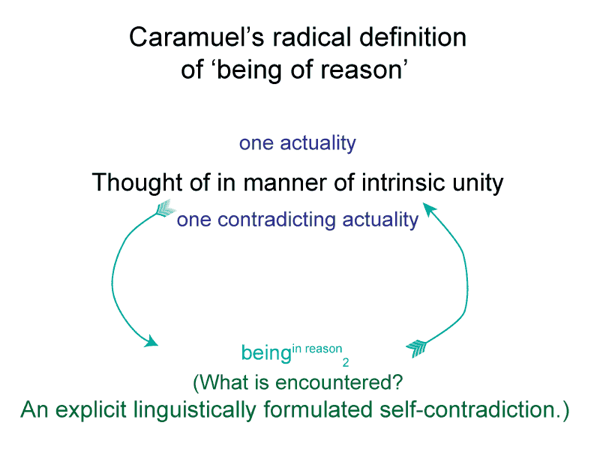
The self-contradiction is also a being of reason. A good example is a large human rat.
0069 If I encountered a large human rat, I would be surprised, since large human rats do not exist. Yet, if I encountered something large, like a raccoon, scurrying inside a dumpster, I could mistake it as both a rat and a human. In this way, self-contradictions may exist as beings of reason.
SN3: Every being is either a real being or being of reason, but no being is both.
0070 The raccoon (neither human nor rat, but easily mistaken for both) is a beingitself2a.
The human rat is a beingin_reason2a.
This suggests the following conclusion. Since self-contradictions, negations and privations are always beings of reason, they can never exist as beingsthemselves.
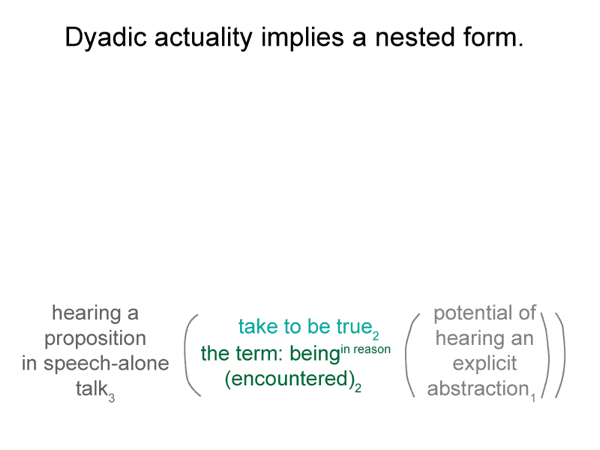
This lesson is plainly portrayed in the content-level actuality in the bare-bones schema for implicit abstraction. The first element of the content-level of actuality is a real being. A second element must be present, according to the nature of secondness. This second element somehow accounts for the actuality of the encountered real being.
0071 At this point, I have a general idea of why schoolmen postulated beings of reason in their various inquiries. After all, when looking at beings, there are all sorts of reversals, losses, confounding things and dependencies. These are grasped as negations, privations, self-contradictions and relations.
What does it mean for one of these nonbeings to fill the emptiness of the second element of the content-level actuality? It becomes a beingin reason. It is taken to be real, even though it does not exist.
SN4: Beings of reason are – in the analogical sense of the word “are”.
SN7: A being of reason is regarded as a being, even though it has no being in itself.
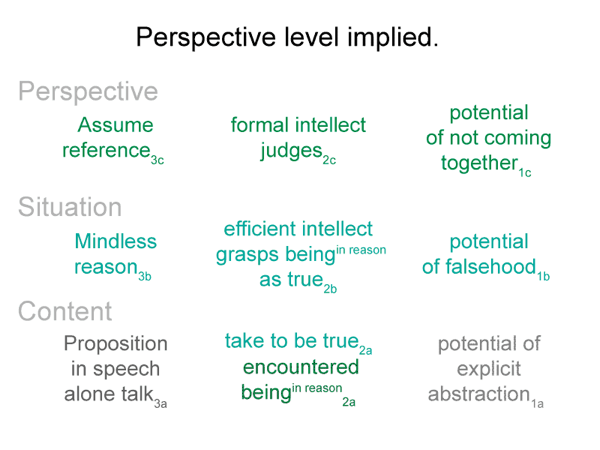
0072 The efficient intellect2b is a real being. It ascribes being (existence) to the beingin_reason2a. “Object” is another word for beingin_reason. The efficient intellect2b grasps an object2a. It makes the object part of its subjective — reflexive — reckoning.
The object2a goes with the content-level nested form. The efficient intellect2b belongs to the situation-level nested form. The content-level normal context3a is what is happening3a. The situation-level normal context3b is situational reason3b.
0073 SN6: A beingin_reason2a has only objective being in the intellect2b.
SN5: Three different relations to reason are indicated by the expression ‘being of reason’. These are (1) effective, (2) subjective and (3) objective.
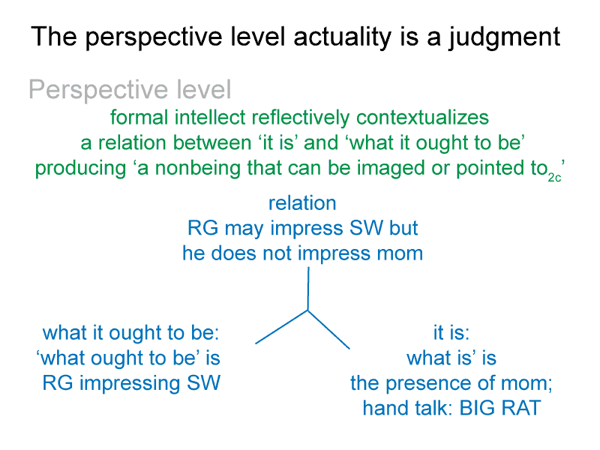
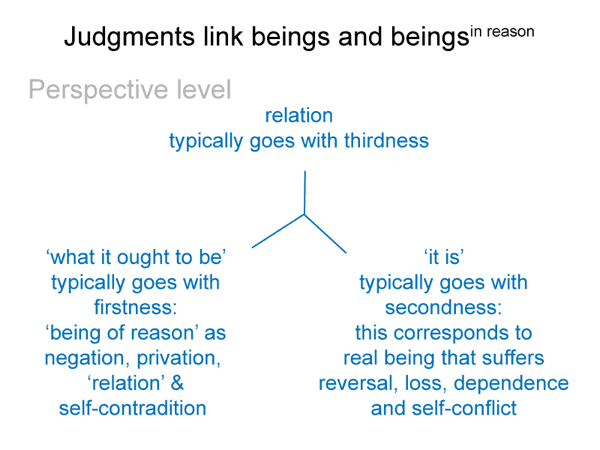
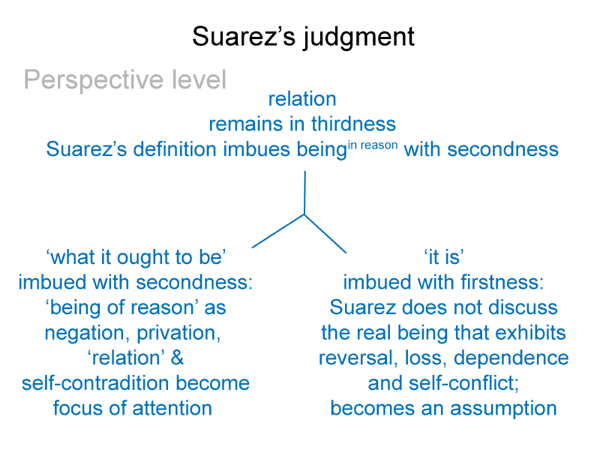
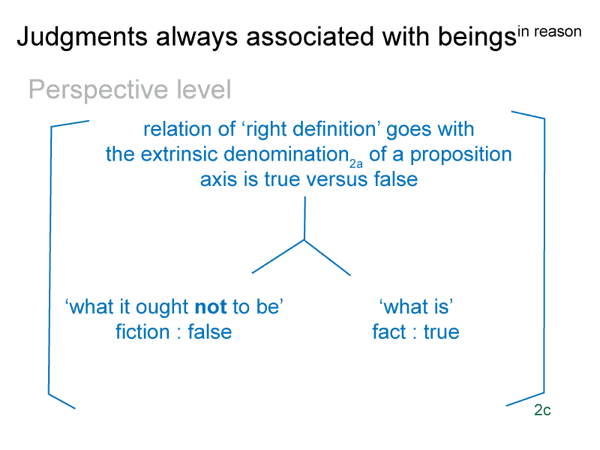
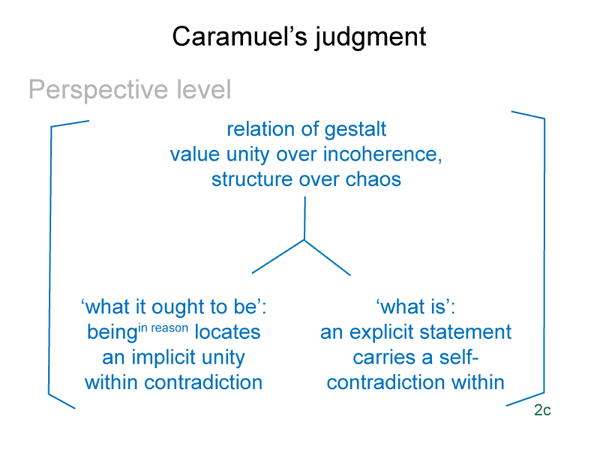
This is a minimum, since the bare-bones schema offers one more. (4) A perspective-level actuality2c draws into relation the ‘what it is’ of the encountered being2a and the ‘what it ought to be’ of the beingin reason2a. Since the intellect, the seat of reason, is involved, Suarez predicted that this relation would generate a comparison.
SN9: Beings of reason and real beings are related by an analogy of proportionality.
0074 Finally, Suarez suggests why ‘beings of reason’ are necessary.
SN8a: ‘Beings of reason’ are necessary to know nonbeing.
Nonbeing is more than nothing. According to the bare-bones schema, the beingin reason2a is an actual object that the efficient intellect grasps.
0075 SN3b: ‘Beings of reason’ are needed to know things relatively in comparison to other things.
This reminds me of the perspective-level nested form. The actuality that puts the efficient intellect into perspective2c is a judgment that weighs ‘what it is’ against ‘what it ought to be’2c. This judgment2c grounds reckoning2b. It addresses the question: On what basis would the efficient intellect2b grasp a real nonbeing2a in response to encountering a real being2a?
0076 SN3c: ‘Beings of reason’ are required to explain the capacity of our intellect to think of self-contradictory beings (as well as negations, privations and relations).
0077 This matches the situation-level nested form. The efficient intellect reckons2b. Once a real being is encountered, the efficient intellect reckons that there must be a second element to the dyad in the content-level actuality. This must be an innate reckoning, because the efficient intellect does exactly what needs to be done. It2b projects an object2a — a beingin_reason2a — into the emptiness of the second element in the realm of actuality2a.
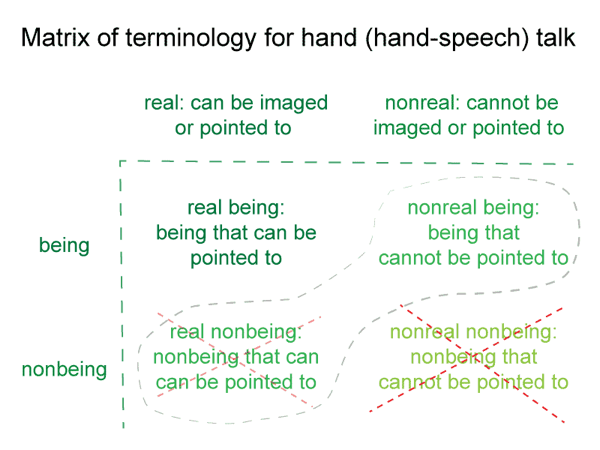
The projected beingin_reason is thought of in the manner of being (existence). This is why humans regard negations, privations, relations and self-contradictions as real, even though they cannot be seen, heard, touched, tasted or smelled.